HÓA HỌC NGÀY NAY – Ứng dụng enzyme trong ngành công nghiệp
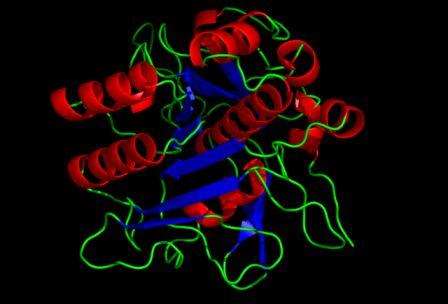 Phân loại protease và subtilisin
Phân loại protease và subtilisin
Protease hay peptidase ( EC. 3.4. -. – ) là nhóm enzyme thủy phân có năng lực cắt mối link peptide ( – CO ~ NH – ) trong những phân tử polypeptide, protein và một số ít cơ chất khác tương tự như thành những amino acid tự do hoặc những peptide phân tử thấp ( Hình 1.1 ). Trong mạng lưới hệ thống mạng lưới hệ thống danh pháp MEROPS ( www.merops.ac.uk ) phân loại protease, subtilisin ( EC. 3.4.21. 62 ) thuộc phân nhánh SB của nhóm serine protease ( EC. 3.4.21. – ) với điểm đặc trưng là sử dụng đơn phân serine ( Ser ) nằm trong TT xúc tác phản ứng của enzyme. Ngoài subtilisin, trong phân nhánh SB còn có những protease tương quan đến quy trình giải quyết và xử lý tiền hormone như KEX2 protease, furin, và PC2. Hiện nay, người ta đã xác lập được 97 loại substilisin từ 1659 trình tự trong đó có 16 substilisin đã được thiết kế xây dựng cấu trúc khoảng trống .
Những protease trong họ S8A này đều có một domain bảo thủ với 7 nếp gấp β được phủ bọc bởi 9 vòng xoắn α. Trung tâm hoạt động giải trí của enzyme này là bộ 3 amino acid truyền thống lịch sử của serine proease theo thứ tự là Asp-His-Ser ( Hình 1.2. Bộ 3 xúc tác Asp-His-Ser đặc trưng của serine protease ) .
Cơ chế xúc tác của subtilisin
Nhóm serine protease là nhóm peptidase lớn nhất và được phát hiện ở mọi giới sinh vật như eukaryote, prokaryote, archaea và virus. Những enzyme này đều có chung một chính sách xúc tác phản ứng thủy phân trải qua hai bước chính ( Barrett, 1994 ) :
· Bước 1, acyl hóa : hình thành link cộng hóa trị giữa nhóm – OH của serine với nguyên tử cácbon trong nhóm cácboxyl của phân tử cơ chất nhờ có tương hỗ của nhóm imidazole từ histidine. Kết quả phản ứng này là tạo ra một hợp chất trung gian và một ion imidazolium ( phản ứng cộng ). Hợp chất trung gian không bền này nhanh gọn bị thủy phân thành một acyl-enzyme, vòng imidazole và một amine ( phản ứng khử ) ( Fastrez and Fersht, 1973 ) .
· Bước 2, khử acyl hóa : phức hệ acyl – enzyme bị thủy phân bởi phân tử H2O theo chiều ngược lại của bước một. Trong đó, nhóm imidazole chuyển proton của gốc – OH từ serine cho nhóm amine để tái sinh lại enzyme ( Hình 1.2 ) .
Một số loại subtilisin
Subtilisin là một nhóm enzyme protease kiềm ( serine protease ) với 2 đại diện thay mặt được nghiên cứu và điều tra nhiều nhất là subtilisin Carlsberg và subtilisin BPN ” .
· Subtilisin Carlsberg. Enzyme này được tìm thấy khi nuôi cấy B. licheniformis vào năm 1947 bởi Linderstrom, Lang và Ottesen tại phòng thí nghiệm Carlsberg ( Phadarate et al., 1997 ). Nó được thu lần tiên phong ở dạng kết tinh vào năm 1952 và từ đó đến nay, subtilisin là protease vi sinh vật công nghiệp quan trọng nhất, được sử dụng nhiều trong sản xuất những chất tẩy rửa .
Hoạt tính của enzyme Carlsberg bị ngưng trệ bởi những chất phản ứng, 3,4 – dichloroisocoumarin ( 3,4 – DCI ), L-3-carboxytrans 2,3 – epoxypropyl-leucylamido ( 4 – guanidine ) butane ( E. 64 ), di-isopropylfluorophosphate ( DFP ), phenylmethyl-sulfonyl fluoride ( PMSF ) và tosyl-L-lysine chloromethyl ketone ( TLCK ). Một số khác lại bị ngưng trệ bởi thuốc thử thiol ví dụ như p-chloromercuribenzoate ( PCMB ), đặc trưng so với toàn bộ những serine protease ( Govind et al., 1981 ). Subtilisin Carlsberg có tính đặc hiệu rộng, thủy phân nhiều loại link peptide cũng như những link ester, nhất là những link được tạo thành từ những amino acid thơm. Dưới tính năng của enzyme này, một số ít triglyceride cũng hoàn toàn có thể bị thủy phân như tripropionin và tributyrin. Subtilisin Carlsberg thủy phân từ 30 đến 40 % link peptide của casein. Giống như những serine protease khác, Ca2 + không ảnh hưởng tác động đến độ bền của enzyme này, có pH tối ưu là 8,0 – 9,0 và bền trong khoanh vùng phạm vi pH rộng từ 5,0 đến 11,0 ( Zuidweg et al., 1972 ) .
· Subtilisin BPN ”. Enzyme này đã được làm sạch và thu được ở trạng thái kết tinh từ chế phẩm công nghiệp BPN ” ( bacterial protease Nagase ). Từ chế phẩm BP Novo ( bacterial protease Novo ) đã tách được enzyme có thành phần amino acid tựa như subtilisin Novo ( Forgarty, 1983 ). Subtilisin BPN ” được tổng hợp từ những loài B. amyloliquefaciens, B. subtilis và B. stearothermophilus ( Zuidweg et al., 1972 ). Subtilisin BPN ” và subtilisin Carlberg khác nhau 58 amino acid nhưng C và pH 10 ( Smith et al., ° chúng hoạt động giải trí tối ưu ở cùng một điều kiện kèm theo là 60 1968 ; Phadarate et al., 1997 ; Siezen and Leunissen, 1997 ). Khác với subtilisin Carlberg độ bền của subtilisin BPN ” lại nhờ vào vào Ca2 +. Subtilisin BPN ” còn là một enzyme ngoại bào nên hoàn toàn có thể thuận tiện tịch thu và tinh sạch hơn những enzyme nội bào, do vậy việc ứng dụng enzyme này ngày một phổ cập .
Subtilisin BPN ” là một serine protease ngoại bào và là một lớp enzyme đại diện thay mặt lớn nhất của serine protease có cấu trúc đặc trưng là α / β / α và TT hoạt động giải trí gồm Ser221, His64, Asp32 ( Hình 1.3 ). Cấu trúc khoảng trống ba chiều của chúng vững chắc trong khoảng chừng pH rộng và ngay cả khi xuất hiện của dung môi hữu cơ ( Siezen and Leunissen, 1997 ; Thomas et al., 1999 ) .
Subtilisin BPN ” là một pre-pro-enzyme ( Wells et al., 1983 ). Trình tự pre – là một chuỗi gồm 30 amino acid có tính năng như một chuỗi peptide tín hiệu trong quy trình tiết subtilisin ra môi trường tự nhiên bên ngoài. Trình tự pro – gồm 77 amino acid được nối với subtilisin chỉ đến khi quy trình gấp nếp kết thúc và sau đó chúng tự động hóa cắt bỏ khỏi phân tử subtilisin trưởng thành. Chính cho nên vì thế chuỗi peptide gồm 77 amino acid này được ví như một ‘ foldase ’ xúc tác cho quy trình gấp nếp của phân tử enzyme. Phân tử subtilisin trưởng thành gồm 275 amino acid và có khối lượng phân tử là 27,5 kDa ( Ikemura and Inouye, 1988 ; Ohta and Inouye, 1990 ). Một số subtilisin khác. Ngoài hai nhóm subtilisin lớn kể trên còn có 1 số ít loại khác như : subtilisin E, subtilisin NAT, Tk-subtilisin ( từ T. kodakaraensis KOD1 ). Sutilisin E và NAT đều được tổng hợp từ B. subtilis và trình tự gene mã hóa cho chúng cũng đã được xác lập ( Stahl and Ferrari, 1984 ; Nakamura et al., 1992 ). Việc nghiên cứu và phân tích trình tự nucleotid cho thấy sutilisin NAT có độ tương đương 99,5 % với subtilisin E, 86 % với subtilisin BPN ” và 72 % với subtilisin Carlsberg ( Tetsumei et al., 2001 ). Tk-subtilisin được tổng hợp từ T. kodakaraensis KOD1 .
Ứng dụng của subtilisin
Với sự phong phú về chủng loại, subtilisin đã và đang được điều tra và nghiên cứu và ứng dụng trong nhiều nghành công nghiệp với những tính năng tương thích. Rất nhiều loại sản phẩm thương mại của subtilisin được đưa ra thị trường, chúng hầu hết được sử dụng trong ngành công nghiệp những chất tẩy rửa, thuộc da và cả trong y dược học do subtilisin có năng lực hoạt động giải trí trong phổ pH rộng và nhiệt độ cao .
Bảng 1.1 Một số thương phẩm subtilisin trên thị trường ( Rao et al., 1998 ; Gupta et al., 2002 ). Lĩnh vực ứng dụng Tên thương phẩm / Hãng Nguồn gốc Bột giặt và chất tẩy rửa BIOTEX ( Alcalasease ) ( Novo Industry A / S ) B. licheniformis Bioprase ( Nagase Biochemicals, Nhật Bản ) B. subtilis Esperase, Savinase T ( Novo Industry ) Bacillus spp .
Sữa tắm Kem tắm làm trắng da NA ( Công ty Trách Nhiệm Hữu Hạn Sản xuất và TM Nhân Lộc ) B. subtilis Công nghiệp thuộc da Aquaderm, NUE, Pyrase ( Novo Nordisk ) Công nghiệp thực phẩm GRAS B. subtilis và Endothia parasitica HT-proteolytic ( Solvay Enzymes, GerSuby ) B. subtilis Công nghiệp dược phẩm Luizym, Nortase Aspergillus oryzae Bioprase concentrate ( Nagase Biochemicals, Nhật Bản ) B. subtilis Đang nghiên cứu và điều tra Cryst, alkaline protease ( Nagase Biochemical, Nhật Bản ) B. subtilis
Chất tẩy rửa
Protease là một trong những thành phần không hề thiếu trong tổng thể những loại chất tẩy rửa, từ chất tẩy rửa dùng trong mái ấm gia đình đến những chất làm sạch kính hoặc răng giả và kem đánh răng. Việc ứng dụng enzyme vào những chất tẩy rửa nhiều nhất là trong bột giặt. Các protease thích hợp để bổ trợ vào chất tẩy rửa thường có tính đặc hiệu cơ chất rộng để thuận tiện vô hiệu những vết bẩn do thức ăn, máu và những chất do khung hình con người tiết ra. Một tiêu chuẩn quan trọng khác của những protease dùng trong chất tẩy rửa là hoạt động giải trí tốt trong điều kiện kèm theo nhiệt độ và pH cao cũng như phải thích hợp với những tác nhân oxy hóa và những chất ngưng trệ có trong thành phần của chất tẩy rửa. Và tham số đóng vai trò chìa khóa cho việc bổ trợ protease nào vào chất tẩy rửa là pI của chúng. Một protease tương thích khi pI của nó trùng với pH của dung dịch chất tẩy rửa. Subtilisin cung ứng được không thiếu những nhu yếu khắc nghiệt trên .
Chất tẩy rửa tiên phong có chứa enzyme vi trùng được sản xuất vào năm 1956 với tên BIO-40. Đến năm 1963, Novo Industry A / S đã ra mắt alcalase dưới tên thương mại là BIOTEX được chiết xuất từ B. licheniformis. Và đến gần đây, toàn bộ những protease bổ trợ vào chất tẩy dùng trên thị trường đều là serine protease được sản xuất từ những chủng Bacillus ( Rao et al., 1998 ; Thangam and S., 2002 ), và hầu hết là từ B. subtilis. Trên quốc tế, mỗi năm người ta đã sử dụng 89 % enzyme này cho ngành công nghiệp tẩy rửa. Trong đó hai công ty lớn là Novo Nordisk và Genencor Internatinal mỗi năm đã cung ứng cho toàn thế giới hơn 95 % lượng enzyme protease ( Gupta et al., 2002 ) .
Công nghiệp thuộc da
Quá trình chế biến da gồm có một số ít quy trình như ngâm ướt, tẩy lông, làm mềm da và thuộc da. Thông thường những giải pháp thuộc da thường dùng những hóa chất ô nhiễm như natri sulfide, làm ảnh hưởng tác động rất nghiêm trọng đến môi trường tự nhiên khi nước thải của nhà máy sản xuất này thải ra sông. Việc sử dụng enzyme để thay thế sửa chữa những hóa chất đã rất thành công xuất sắc trong việc nâng cao chất lượng da và làm giảm ô nhiễm môi trường tự nhiên. Protein là một thành phần cơ bản của da và lông nên protease đã được sử dụng để thủy phân 1 số ít thành phần phi collagen của da và vô hiệu những protein phi fibrin như albumin, globulin trong quy trình thuộc da rất có hiệu suất cao ( Christner, 1996 ; Varela et al., 1997 ; Thangam and S., 2002 ) .
Ứng dụng khác
Do protease kiềm từ Bacillus được tạo thành với lượng lớn, có đặc tính vững chắc, hoạt động giải trí tốt với nhiệt độ và pH cao nên chúng được ứng dụng ở nhiều ngành công nghiệp khác nhau như : giải quyết và xử lý phim X-quang đã qua sử dụng để nhằm mục đích tịch thu bạc ( Fujiwara et al., 1991 ; Ishikawa et al., 1993 ), làm nước mắm cá ( Rebeca et al., 1991 ), làm thức ăn gia súc ( Cheng et al., 1995 ), giải quyết và xử lý chất thải từ động vật hoang dã giáp xác ( Yang et al., 2000 ), giải quyết và xử lý rác thải trong những lò mổ gia cầm ( Dalev, 1994 ) .
Nghiên cứu subtilisin của Bacillus B. subtilis
Theo diễn đạt trong khóa phân loại của Bergey, vi trùng B. subtilis là một loài thuộc chi Bacillus, thuộc họ Bacillacaea. B. subtilis là trực khuẩn Gram dương có size 2-3 x 0,7 – 0,8 µm, nội bào tử ở TT có kích cỡ 1,5 – 1,8 x 0,8 µm. Ở điều kiện kèm theo 100 oC, bào tử của B. subtilis chịu được 180 phút, có tính không thay đổi cao với nhiệt độ thấp và sự khô cạn, ảnh hưởng tác động của hóa chất, tia bức xạ ( Kerovuo et al., 2000 ) .
Năm 1971 lưu lại bước mở màn quan trọng cho những điều tra và nghiên cứu về protease kiềm ở Bacillus. Horikoshi là người tiên phong công bố thu nhận và Markland khám phá cấu trúc cơ bản và đặc thù lý hóa của protease kiềm từ Bacillus ( Horikoshi, 1971 ; Markland and Smith, 1971 ). Sang năm 1972, người ta đã nghiên cứu và điều tra và đưa vào sản xuất trên quy mô lớn những protease kiềm ở một vài loài Bacillus. Đến thập kỷ 80, những điều tra và nghiên cứu về protease kiềm của Bacillus liên tục lan rộng ra và đã tạo ra hàng loạt những loại sản phẩm trên thị trường thương mại. Các vi trùng Bacillus ưa kiềm trở thành “ một quốc tế vi sinh vật mới ” theo “ quan điểm công nghiệp ( Horikoshi and Akiba, 1982 ; Horikoshi, 1996 ) .
Gần đây, người ta đã thành công xuất sắc trong nhân dòng phân tử, giải trình tự nucleotide và bộc lộ gene mã hóa protease kiềm từ nhiều loài Bacillus ( Desmond et al., 1975 ; Harry and Hausinger, 1989 ; Gupta et al., 2002 ). Các vector biểu lộ thường dùng để biểu lộ gene mã hóa subtilisin là : pET22b ( + ), pBS42, pUB110, pHY300PLK, pS1, pUC118, pUC119 …
Cũng như những gene khác, gene mã hóa subtilisin hoàn toàn có thể biểu lộ ở nhiều chủng chủ khác nhau. Các chủng chủ được sử dụng để biểu lộ gene mã hóa subtilisin :
· Escherichia coli những chủng : BL21, C600, HB 101, JM 109
· Bacillus những chủng : subtilis I-168, DB110, stearothermophilus
· Nấm : Saccharomyces serevisiae, Aspergillus oryzae, Streptococcus cremoris, Streptomyces lividans
Trong đó các chủng B. subtilis được sử dụng nhiều nhất vì đã được nghiên cứu tương đối đầy đủ về đặc tính sinh lý, sinh hóa và di truyền với toàn bộ genome đã được xác định. So với E. coli, B. subtilis có ưu điểm là vi khuẩn Gram dương không gây bệnh và không chứa độc tố. Tuy nhiên chúng có nhược điểm là plasmid tái tổ hợp chứa DNA ngoại lai thiếu bền vững nên việc biểu hiện trên B. subtilis thường yêu cầu tái tổ hợp vào nhiễm sắc thể. Do đó, để khảo sát đặc tính một enzyme mới người ta thường lựa chọn biểu hiện trên E. coli trước. Sau đó, khi sản xuất protein lượng lớn thì chuyển sang hệ thống biểu hiện của B. subtilis.
Xem thêm: Quỹ đầu tư phát triển là gì?
Hoahocngaynay.com
Nguồn Viện Nghiên cứu Da Giầy
Chia sẻ :
Source: https://laodongdongnai.vn
Category: Doanh Nghiệp





